|
Immunology Laboratory |
Basic Concepts |
| |
When a foreign antigen is
introduced into an animal, the animal will respond immunologically to
it. Innate (natural) responses occur as many times the foreign
agent is encountered and lacks immunological memory, whereas acquired
(adaptive) responses improve on repeated exposure to the antigen. The
immunological response may be of two different types: cell-mediated
or humoral.
|
|
Organization of the Immune System |
|
The cells involved in the immune response
are effectively organized into tissues and organs. The
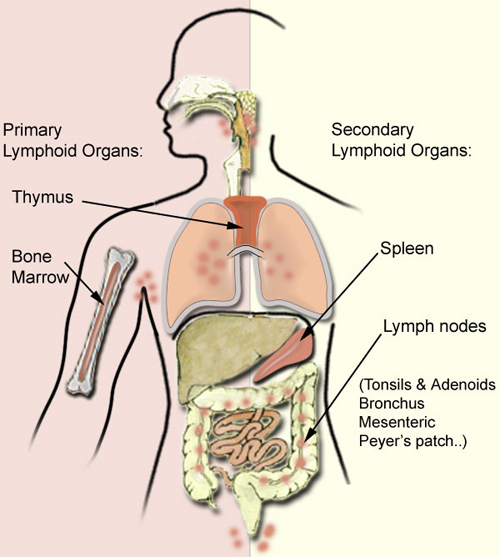
major lymphoid organs are classified into either primary or secondary.
Primary lymphoid organs (thymus
and bone marrow) are the major sites
of lymphocyte development (lymphopoiesis). Their function is to
produce a large repertoire of reactive cells (they acquire their
repertoire of specific antigen receptors) and to eliminate self-reacting
cells (cells with receptors for autoantigens are mostly eliminated).
Secondary lymphoid organs (spleen, lymph
nodes, mucosal associated lymphoid tissue) provide the
environment for the proliferation and maturation of cells
involved in the adaptive immune response, for filtering and trapping
antigens. They also provide the environment for cell-cell
interaction and cytokine-cell interaction. |
|
|
|
|
|
Cells of the Immune System |
|
Immune responses are mediated by a number of
cells and by the soluble molecules they secrete. |
|
|
|
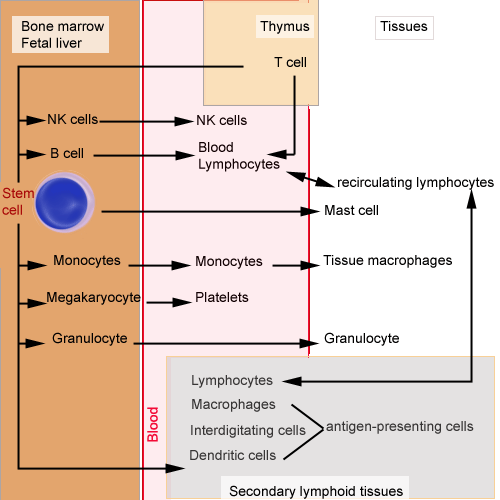
Origin of cells of
the immune system: all the cells shown above arise from the
hemapoietic stem cell. Platelets are released into the circulation.
Granulocytes and monocytes pass from the circulation into the tissues.
Mast cells are evidenced in all tissues. B cells mature in the primary
lymphoid organs (bone marrow...). T cells mature in the thymus
(primary lymphoid organs). Lymphocytes leave primary lymphoid organs and recirculate through secondary
lymphoid tissues. Interdigitating and dendritic cells act as
antigen-presenting cells in secondary lymphoid tissues. |
|
Function of the cells of
the immune system: |
|
The innate responses use phagocytic cells
(neutrophils, monocytes, and macrophages), cells which release
inflammatory mediators (basophils, mast cells, and eosinophils), and
natural killer cells. The molecular components of the innate responses
comprise complement, acute-phase proteins, and cytokines such as the
interferons. |
Acquired responses involve the proliferation
of antigen-specific B and T cells, occurring when their surface
receptors bind to the antigen. Specialized cells (antigen-presenting
cells) display the antigen to T lymphocytes and collaborate with them in
the response to the antigen.
B cells secrete immunoglobulins (antigen specific antibodies)
responsible for eliminating extracellular microorganisms or foreign
agents. Innate and acquired responses usually work together. |
|
Major Histocompatibility Complex (MHC): this
gene complex was first identified when it was observed that
histocompatibility depended on the donor and recipient sharing the same
haplotype. The molecules which determine graft rejection are a limited
group: Class I and Class II of cell-surface structures. The molecules
which present antigens to T cells are mostly encoded within the MHC. All
nucleated cells of the body express MHC Class I. By contrast, MHC Class
II molecules are used by antigen presenting cells to present antigens to
helper T cells, so cells expressing Class II are smaller in numbers than
those expressing Class I. |
|
Antigen presenting cells, in a
lymphoid organ, include macrophages, dendritic cells and B-cells. |
|
|
|
Mononuclear phagocytes: most important group
of long-lived phagocytic cells. These cells are bone marrow-derived and
their function is to engulf particles, internalize them and destroy
them. |
|
Dendritic
cells which are found in the
T-cell areas of lymph node and spleen are the most effective cells for
the initial activation of naive T-cells. |
|
B-cells: bone marrow-derived. Each B cell is
genetically programmed to encode a surface receptor specific for a
particular antigen. Upon recognition of the antigen, B cells multiply
and differentiate into plasma cells, which produce large amounts of the
receptor molecule in a soluble form that can be secreted (antibody). |
 |
|
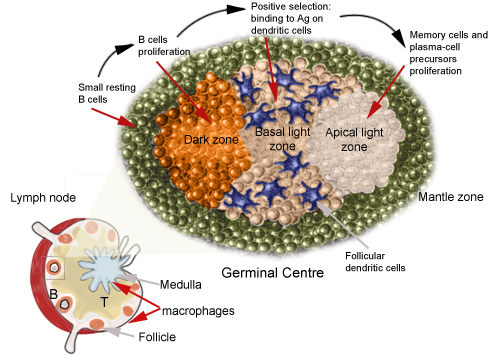
The germinal centre:
with the initiation of the acquired immune response, germinal centres
form in the secondary lymphoid tissues, where all the antigen-specific
and antigen-presenting cells can interact. |
|
|
|
|
|
Effector cells: T-cells and B
lymphocytes. |
T cells: there are several types of T
cells with a variety of functions. They originate from bone marrow stem
cells and require further differentiation in the thymus where they
migrate. T cell maturation requires a number of cell interactions:
T cells express in an orderly fashion certain markers or cell surface
proteins. The nomenclature used to refer to cell surface molecules,
characterized on the basis of their reactivity to monoclonal antibodies,
follows the CD (for "cluster of differentiation") numbering. The
CD4+
T cells, cytokine-secreting helper cells can be divided into two major
types:
- Type I helper T cells TH1 which
secrete Interleukin (IL) 2 and interferon (IFN)
g.
- Type II helper T cells TH2 which
secrete IL4 and 5.
The production of cytokines by TH1
facilitates cell-mediated immunity (activation of macrophages and T-cell
mediated cytotoxicity). TH2 cells help B cells produce antibodies. Class
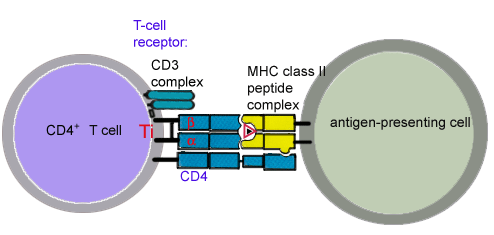
II MHC on antigen presenting cells interact with CD4 on T cells.
Peptides which bind to MHC Class II come from proteins which have been
internalized by the cell and then degraded.
CD8+ T cells play a role in the elimination of virally infected
cells. The infected cell marks itself as a target for the cytotoxic T
cell by displaying peptides derived from intracellular viral protein on
its surface. Viral proteins are bound to peptide-binding regions of
class I MHC molecules. Peptides which bind to MHC Class I molecules come
from proteins synthesized within the cell (which are broken down) and
transported to the endoplasmic reticulum. |
B cells: the earliest cells which
develop are called B1 cells; they express CD5 cell-surface molecule and
are the source of "natural antibodies", which are IgM antibodies and are
frequently polyreactive (recognize different antigens, pathogens and
autoantigens). Natural antibodies have a relative low affinity. Most B
cells lack the CD5 molecule, they develop later and are called B2 cells.
Mature B2 cells coexpress IgM and IgD antibodies on their cell surface.
The genes encoding B-cell receptors undergo a process of somatic
hypermutation; the final stages of differentiation of B2 cells into
antibody secreting plasma cells occurs within the germinal centres of
secondary lymphoid tissues.
To elicit a strong antibody response, B cells require:
-
Antigen
-
T cells for direct contact (usually TH2
cells)
-
Soluble cytokines (e.g. IL4 + IL13, INF-γ or
IL10)
-
Certain adhesion molecules
|
|
Clonal selection involves the proliferation
of cells which recognize a specific antigen: each B cell is programmed
to make just one antibody specificity, located on its surface as an antigen
receptor. Antigen binds to only those B cells with the appropriate
surface receptor; these cells are stimulated to proliferate and mature
into antibody producing cells. |
 |
 |
|
Functions of Antibody |
|
The primary function of the antibody is to
bind the antigen; the antibody directs specificity towards the antigen.
By doing do, it also arms the killer cells and activates complement,
processes that eliminate foreign organisms, which the antibody by itself
cannot do. Of the different classes of antibodies, two will be discussed
briefly: |
|
IgM class: is the predominant antibody (it
is the first to appear) in primary immune responses and is associated
with the immune responses to antigenically complex, blood -borne agents.
Once bound to the antigen, it is a powerful activator of the classical
pathway complement: a single molecule of bound IgM is able to initiate
the cascade because of the adjacent positioning of the Fc regions.
|
|
IgG class: is the most important class of
immunoglobulin in secondary immune responses |
|
|
|
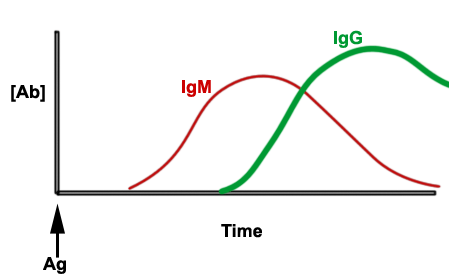
The exposure to foreign antigen yields a
biphasic response. The first phase is associated with production of IgM,
followed by production of IgG. The second phase is characterized by a
reduction of IgM followed by an increase of IgG. The antigen will select
and expand a clone of effector B cells which will develop into plasma
cells and produce antibodies. |
|
Complement system |
|
The complement system consists of about 20+
serum molecules (some of them being proteases) constituting nearly 10%
of the total serum proteins and forming one of the major defence systems
of the body. The complement system is activated by the Ag-Ab complex.
The principle functions are the initiation of: |
|
chemotactic factors:
polymorphs and macrophages have specific receptors for small complement
fragments generated during complement activation. The fragments diffuse
away from the site of activation and stimulate chemotaxis, the way
chemokines do. |
|
vasodilation
factors: some of the protein fragments of the complement
system (C5a) causes degranulation of mast cells and basophils with
release of histamine and other vasoactive mediators. Consequently, there
are indirect effects on blood vessels, vasodilation and increased
permeability of capillaries. |
|
factors that
increase phagocytosis: phagocytic cells carrying receptors
for the complement components are then able to bind to the foreign
particle (opsonization), and this triggers phagocytosis and cell
activation. |
|
membrane attack
complex: the final step in complement activation brings about
the assembly of a membrane attack complex, which can insert itself into
lipid bilayers, causing the lysis of the foreign body. |
|
The lytic pathway: |
|
|
|
 |
|
Click here to continue with the topic of Elisa |
)